| Laboratory of Cornelius Krasel | ||||||||
| Research | Teaching | Publications | People | |||||
Cell Biology of G-protein-coupled receptors (GPCRs)
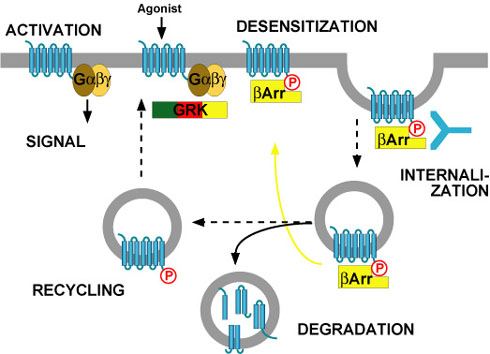
As shown in the scheme,
desensitisation, internalisation and recycling
of G-protein-coupled receptors are intimately connected. The whole process
is initiated by binding of an agonist to the receptor, which triggers a
signal (usually via heterotrimeric G-proteins). However, agonist binding
also converts the receptor into a substrate for a family of kinases, the
G-protein-coupled receptor kinases (GRKs). These kinases phosphorylate
only agonist-activated receptors. Subsequently, the phosphorylated receptor
becomes a binding partner for arrestins. Arrestins are normally cytosolic
proteins, but they recognise agonist-activated, phosphorylated receptors
and bind them. This binding makes the receptor inacessible for G-proteins
(i.e. the arrestin-bound receptor is desensitised), and it targets the
receptor for internalisation. This is because arrestins do not only bind
receptors, but they also bind components of clathrin-coated pits. Thus,
arrestin-bound receptors move into clathrin-coated pits and are then
internalised.
We are using a variety of methods to investigate these processes. We are able to purify most of the participating proteins from either bacteria or insect cells. Thereby we can investigate regulation of these proteins in vitro. Recently we have produced arrestin2 in a new crystal form (PDB entry 2WTR). We also look at binding between proteins in vitro. However, we also perform a range of experiments in living cells. For example, we can determine the internalisation of receptors by ligand binding. We also look at translocation of GFP-tagged proteins by live-cell confocal microscopy (see example below). Furthermore, we investigate protein-protein interactions in single living cells using fluorescence resonance energy transfer (FRET) between CFP- and YFP-tagged interaction partners. Finally, we determine the affinity of GPCR-binding proteins by two-color fluorescence recovery after photobleaching (FRAP).
At the moment, we are particularly interested in the interaction mechanism of arrestins with G-protein-coupled receptors. To address these questions, we use the methods mentioned above and complement them with more classical protein biochemistry. Recently, we could show that the co-internalization of arrestins with some GPCRs can be modulated by altering the affinity of GPCR-arrestin interaction. We could also demonstrate that receptor recycling to the plasma membrane after agonist removal is not related to co-internalization of arrestins with receptors, as previously postulated. We also collaborate with
other laboratories working on the interaction of G-protein-coupled receptors
with GRKs and arrestins. These collaborations currently focus on the
μ opioid receptor, the apelin receptor and the receptors for glucagon-like
peptide 1 (GLP1) and glucose-dependent insulinotropic polypeptide (GIP).
|
 The effects of drugs often diminish over time. This "desensitization"
phenomenon forms the basis of my research. Specifically, we are studying the
desensitization of G-protein-coupled receptors. These proteins are usually
localized within the plasma membrane where they sense a wide variety of
extracellular signals and transmit them to the cell interior. They are targets
for more than 30% of all prescription drugs. Therefore, understanding the
mechanisms of their desensitization - and possibly preventing its occurrence -
is of paramount interest.
The effects of drugs often diminish over time. This "desensitization"
phenomenon forms the basis of my research. Specifically, we are studying the
desensitization of G-protein-coupled receptors. These proteins are usually
localized within the plasma membrane where they sense a wide variety of
extracellular signals and transmit them to the cell interior. They are targets
for more than 30% of all prescription drugs. Therefore, understanding the
mechanisms of their desensitization - and possibly preventing its occurrence -
is of paramount interest.